Models of synaptic conductance II
NOTE In this set of articles I would like to cover three simple models of synaptic conductance which are typically used in spiking networks. While people are quite familiar with their general logic, the exact mathematical details are often overlooked or simply swept under the rug because they are deemed too simple. I will take on this all-too-simple task, and talk about the mathematical ideas behind these basic models.
In the first article of this series we discussed synapses which activate instantaneously. These synapses are easy to program, and offer an excellent approximation when one is interested in phenomena occurring at timescales larger than the activation dynamics. However, once we go down to about
Here,
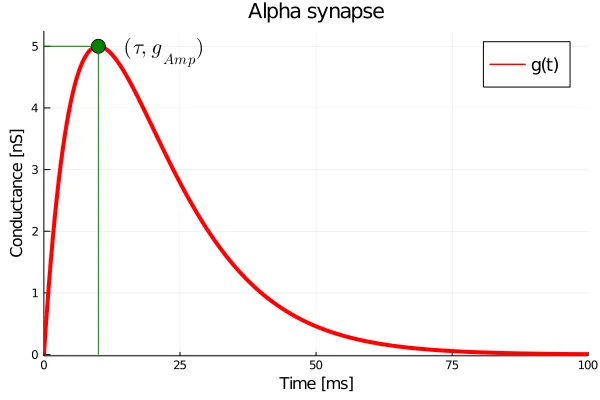
To gain a better intuition about the behaviour of the alpha synapse, let us try to formulate its equations of motion. A first attempt makes it clear that a single state variable does not suffice, i.e., it is not as ‘memoryless’ as the ISED synapse. However, after some manipulation one can arrive at:
We have introduced a second variable
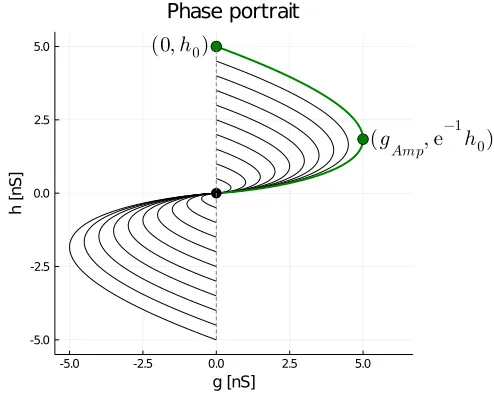
There are a few things to note here. First, I have only plotted the first and third quadrants as synapses cannot be both excitatory and inhibitory. Second, the sense of all trajectories is towards the origin (black circle) - this is not evident from the plot. For most trajectories, the conductance (i.e., the abscissa) grows, attains a maximum amplitude, and then decays to zero. Furthermore, given a trajectory
Now for the spiking. What do you think happens when the synapse receives another pre-synaptic spike while the conductance is still non-zero? Ideally, one would want the system to not only reset, but to also remember the impact from the previous inputs. In ISED synapses, this memory is not needed as the impact is instantaneous. However, in alpha synapses this memory is crucial, and can be implemented by a simple extension of equation
Here, the
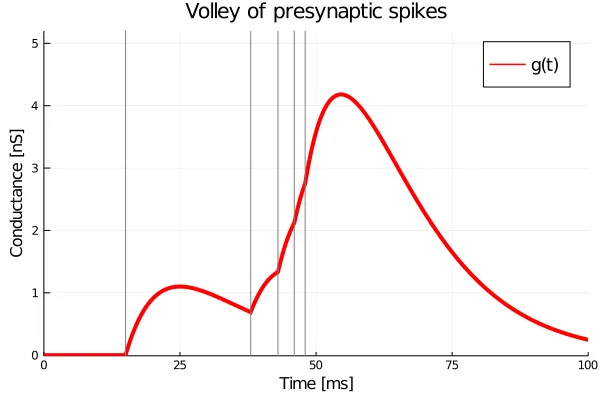
Here, the arrival times of the input spikes are shown using grey vertical lines, and each input spike has
The alpha synapse offers an excellent first approximation of the synaptic activation dynamics, and suffices for most simulations. However, it assumes equivalent time-scales for both the rise and the decay of the impulse response - something that may not be desirable when one wants to model specific transfer functions in detail. In the next and final article of the series, we will look at a second approximation of the synaptic transfer function which tries to address this problem.